De prikkelende postsynaptische potentie is een opwindend potentieel in het postsynaptische membraan van neuronen. De afzonderlijke potentialen worden ruimtelijk en tijdelijk samengevat en kunnen zo een actiepotentiaal creëren. Overdrachtsstoornissen zoals myasthenia gravis of andere myasthenieën verstoren deze processen.
Wat is het prikkelende postsynaptische potentieel?

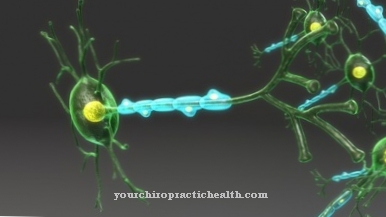
Neuronen zijn van elkaar gescheiden door een opening van 20 tot 30 nm, ook wel bekend als een synaptische opening. Het is de minimale kloof tussen het presynaptische membraangebied van een neuron en het postsynaptische membraangebied van de stroomafwaartse zenuwcel.
Neuronen zenden excitatie uit. Daarom wordt hun synaptische kloof overbrugd door het vrijkomen van biochemische boodschappersubstanties, die ook bekend staan als neurotransmitters. Dit creëert een prikkelende postsynaptische potentiaal op het membraangebied van de stroomafwaartse cel. Het is een lokaal beperkte verandering in het postsynaptische membraanpotentieel. Deze geleidelijke verandering in potentieel triggert een actiepotentiaal in het postsynaptische element. Het prikkelende postsynaptische potentieel maakt deel uit van de neuronale excitatiegeleiding en ontstaat wanneer het stroomafwaartse celmembraan wordt gedepolariseerd.
De opwindende postsynaptische potentialen worden ontvangen en verwerkt door het volgende neuron door zowel ruimtelijk als tijdelijk op te tellen. Wanneer de drempelpotentiaal van de cel wordt overschreden, wordt een nieuw gevormde actiepotentiaal meegevoerd door het axon.
Het tegenovergestelde van het prikkelende postsynaptische potentieel is het remmende postsynaptische potentieel. Dit leidt tot hyperpolarisatie op het postsynaptische membraan, waardoor het activeren van een actiepotentiaal wordt voorkomen.
Functie en taak
Het opwindende postsynaptische potentieel en het remmende postsynaptische potentieel hebben invloed op alle zenuwcellen. Wanneer hun drempelpotentieel wordt overschreden, depolariseren zenuwcellen. Ze reageren op deze depolarisatie door prikkelende neurotransmitters vrij te geven. Een bepaalde hoeveelheid van deze stoffen activeert de zendergevoelige ionenkanalen in het neuron. Deze kanalen zijn doorlaatbaar voor kalium- en natriumionen. Lokale en gegradueerde potentialen in de zin van een prikkelend potentieel depolariseren dus het postsynaptische membraan van het neuron.
Wanneer de membraanpotentiaal intracellulair wordt afgeleid, is de prikkelende postsynaptische potentiaal de depolarisatie van het somamembraan. Deze depolarisatie vindt plaats als gevolg van passieve voortplanting. Er is een opsomming van individuele mogelijkheden. De hoeveelheid afgegeven neurotransmitter en de grootte van de heersende membraanpotentiaal bepalen de omvang van de prikkelende postsynaptische potentiaal. Hoe hoger de pre-depolarisatie van het membraan, hoe lager het prikkelende postsynaptische potentieel.
Als het membraan al boven zijn rustpotentiaal is gedepolariseerd, daalt het postsynaptische prikkelingspotentieel en bereikt onder bepaalde omstandigheden nul. In dit geval wordt het omkeerpotentieel van het prikkelingspotentieel bereikt. Als de pre-depolarisatie nog hoger blijkt te zijn, ontstaat er een potentiaal met een tegengesteld teken. Het prikkelende postsynaptische potentieel is dus niet altijd gelijk te stellen met een depolarisatie. Het beweegt het membraan eerder naar een bepaald evenwichtspotentiaal, dat vaak onder het respectieve rustmembraanpotentiaal blijft.
Hierbij speelt het werk van een complex ionenmechanisme een rol. Met het prikkelende postsynaptische potentieel kan een verhoogde membraanpermeabiliteit voor kalium- en natriumionen worden waargenomen. Aan de andere kant kunnen ook potentialen met verminderde geleidbaarheid voor natrium- en kaliumionen optreden. In deze context wordt aangenomen dat het ionenkanaalmechanisme de trigger is voor het sluiten van alle lekkende kaliumionenkanalen.
Het remmende postsynaptische potentieel is het tegenovergestelde van het prikkelende postsynaptische potentieel. Ook hier verandert de membraanpotentiaal lokaal op het postsynaptische membraan van zenuwcellen. Hyperpolarisatie van het celmembraan treedt op bij de synaps, die het triggeren van actiepotentialen in het kader van het prikkelende postsynaptische potentieel remt. De neurotransmitters bij de remmende synapsen veroorzaken een celreactie. De kanalen van het postsynaptische membraan gaan open en laten kalium- of chloride-ionen passeren. De resulterende uitstroom van kaliumionen en de instroom van chloride-ionen veroorzaakt de lokale hyperpolarisatie in het postsynaptische membraan.
Uw medicatie vindt u hier
➔ Geneesmiddelen tegen spierzwakteZiekten en aandoeningen
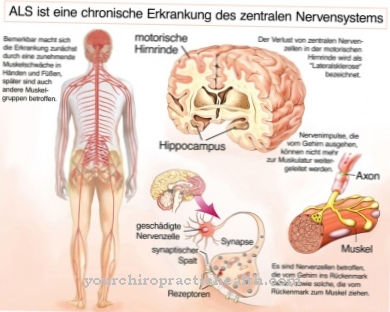
Verschillende ziekten verstoren de communicatie tussen individuele synapsen en dus ook de signaaltransductie bij de chemische synaps. Een voorbeeld is de neuromusculaire ziekte myasthenia gravis, die de eindplaat van de spieren aantast. Het is een auto-immuunziekte met een voorheen onbekende oorzaak. In het geval van de ziekte vormt het lichaam auto-antilichamen tegen het eigen weefsel van het lichaam. Bij spierziekte zijn deze antilichamen gericht tegen het postsynaptische membraan op neuromusculaire eindplaten. Meestal zijn de auto-antilichamen bij deze ziekte acetylcholinereceptor-antilichamen. Ze vallen de nicotine-acetylcholinereceptoren aan op de verbindingspunten tussen zenuwen en spieren. De resulterende immunologische ontsteking vernietigt het lokale weefsel.
Als gevolg hiervan wordt de communicatie tussen zenuw en spier verstoord, aangezien de interactie tussen acetylcholine en zijn receptor wordt bemoeilijkt of zelfs voorkomen door de acetylcholinereceptor-antilichamen. Het actiepotentiaal kan daarom niet meer van de zenuw naar de spier gaan. De spier is daardoor niet meer prikkelbaar.
De som van alle acetylcholinereceptoren wordt verminderd terwijl de receptoren worden vernietigd door de immuunactiviteit. De subsynaptische membranen vallen uiteen en endocytose creëert een autofagosoom. Transportblaasjes versmelten met de autofagsomen en de acetylcholinereceptoren veranderen als gevolg van deze immuunreactie. Met deze wijzigingen verandert de gehele eindplaat van de motor. De synaptische kloof wordt groter. Om deze reden diffundeert acetylcholine uit de synaptische spleet of wordt het gehydrolyseerd zonder te binden aan de receptor.
Andere myasthenieën vertonen vergelijkbare effecten op de synaptische spleet en het prikkelende postsynaptische potentieel.









.jpg)

.jpg)
.jpg)
