De Beëindiging is de laatste fase in DNA-replicatie. Het wordt voorafgegaan door inwijding en verlenging. Een vroegtijdige beëindiging van de replicatie kan resulteren in de expressie van verkorte eiwitten en dus een mutatie.
Wat is de beëindiging?

Tijdens replicatie of reduplicatie wordt het genetische informatiedrager-DNA vermenigvuldigd in individuele cellen. De duplicatie vindt plaats volgens het semi-conservatieve principe en leidt meestal tot een exacte duplicatie van de genetische informatie. Replicatie wordt geactiveerd tijdens de synthesefase, vóór de mitosefase, en vindt dus plaats voordat de celkern zich deelt.
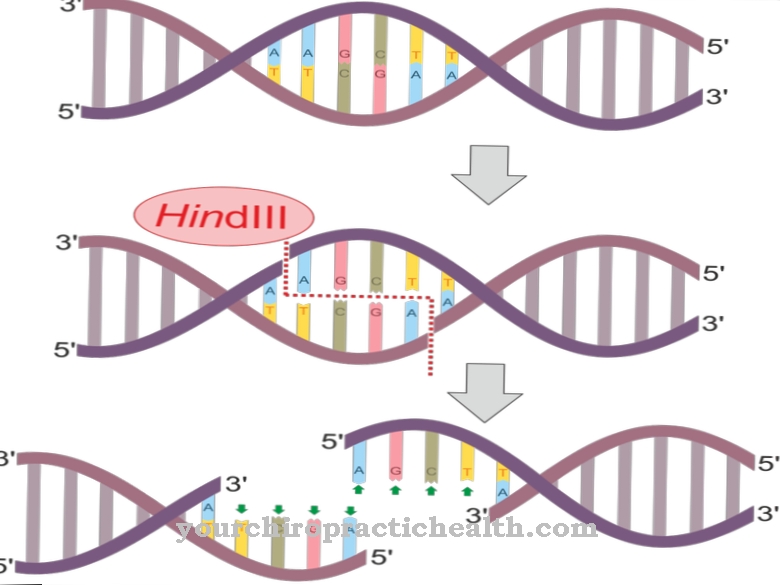
Aan het begin van de replicatie wordt de dubbele DNA-streng gescheiden in enkele strengen, waarop nieuwe complementaire strengen worden gevormd. Elke DNA-streng wordt bepaald door de basesequentie van de tegenoverliggende streng. DNA-replicatie vindt plaats in verschillende fasen. Beëindiging is de derde en dus laatste fase van replicatie. De beëindiging wordt voorafgegaan door initiatie en verlenging.
Een synonieme term voor de uitdrukking van beëindiging in deze context is de aanduiding Beëindigingsfase. Beëindiging betekent hier "afbreken" of "beëindiging". Tijdens de beëindiging raakt de nieuw gevormde mRNA-streng los van het eigenlijke DNA. Het werk van het DNA-polymerase loopt langzaam ten einde. Het beëindigen van DNA-replicatie moet niet worden verward met het beëindigen van replicatie van RNA.
Functie en taak
In de replicatiefase van initiatie vindt primair de regulatie van replicatie plaats. Het startpunt van de replicatie wordt bepaald en er vindt zogenaamde priming plaats. Na de initiatie begint de polymerisatie, waarbij de rekfase plaatsvindt. Het enzym DNA-polymerase scheidt complementaire DNA-strengen in enkele strengen en leest de bases van de enkele strengen een voor een uit. In deze fase vindt semi-discontinue verdubbeling plaats, waaronder een andere fase van priming.
Pas na initiatie en verlenging volgt de beëindigingsfase binnen replicatie. De beëindiging verschilt van levensvorm tot levensvorm. Bij eukaryoten zoals mensen is het DNA gestructureerd in een ring. Het bevat ook terminatiesequenties die overeenkomen met twee verschillende sequenties, die elk relevant zijn voor een replicatievork.
De beëindiging wordt meestal niet geactiveerd door speciale mechanismen. Zodra twee replicatievorken samen lopen of het DNA eindigt, wordt de replicatie op dit punt automatisch beëindigd. De replicatie wordt beëindigd in een automatisch mechanisme.
Beëindigingssequenties zijn besturingselementen. Ze zorgen ervoor dat de replicatiefase op een gecontroleerde manier op een specifiek eindpunt aankomt ondanks de verschillende replicatiesnelheden in de twee replicatievorken. Alle terminatieplaatsen komen overeen met bindingsplaatsen voor het Tus-eiwit, de "terminus-gebruikende stof". Dit eiwit blokkeert de replicatieve helicase DnaB en stopt zo de replicatie.
Bij eukaryoten blijven de gerepliceerde ringstrengen zelfs na replicatie met elkaar verbonden. De verbinding komt overeen met het aansluitpunt. Pas na celdeling worden ze door verschillende processen gescheiden en kunnen ze dus worden verdeeld. De verbinding die tot na de celdeling blijft bestaan, blijkt een rol te spelen bij de gecontroleerde distributie.
Er zijn twee hoofdmechanismen die een rol spelen bij de uiteindelijke scheiding van de DNA-ringen. Enzymen zoals type I en type II topoisomerase zijn betrokken bij de scheiding. Ten slotte herkent een hulpproteïne het stopcodon tijdens de beëindiging. Het polypeptide valt dan van het ribosoom af, aangezien er geen t-RNA met een geschikt anticodon voor het stopcodon beschikbaar is. Het ribosoom valt uiteindelijk uiteen in zijn twee subeenheden.
Ziekten en aandoeningen
Alle processen voor het dupliceren van het genetisch materiaal in de zin van replicatie zijn gecompliceerd en vereisen een hoog verbruik van stoffen en energie in de cel. Om deze reden kunnen spontane replicatiefouten gemakkelijk optreden. Als het genetisch materiaal spontaan verandert of van buitenaf wordt geïnduceerd, spreken we van mutaties.
Replicatiefouten kunnen resulteren in ontbrekende basen, kunnen worden geassocieerd met gewijzigde basen of het resultaat zijn van onjuist basenparen. Bovendien kan het verwijderen en invoegen van enkele of meerdere nucleotiden binnen de twee DNA-strengen leiden tot replicatiefouten. Hetzelfde geldt voor pyrimidine-dimeren, strengbreuken en verknopingsfouten in de DNA-strengen.
Er zijn aparte reparatiemechanismen beschikbaar in het geval van een replicatiefout. Veel van de genoemde fouten worden zoveel mogelijk gecorrigeerd door DNA-polymerase. De replicatienauwkeurigheid is relatief hoog. Het foutenpercentage is slechts één fout per nucleotide, wat te wijten is aan verschillende controlesystemen.
Een controlemechanisme van eukaryote cellen staat bijvoorbeeld bekend als onzin-gemedieerd mRNA-verval, dat ongewenste stopcodons in het mRNA kan herkennen en zo kan voorkomen dat verkorte eiwitten expressie vinden.
Voortijdige stopcodons in het mRNA zijn te wijten aan genmutaties. Zogenaamde onzinmutaties of alternatieve en foutieve splitsingen kunnen resulteren in verkorte eiwitten die worden beïnvloed door functionele verliezen. De controlemechanismen kunnen de fouten niet altijd corrigeren.
De autosomaal recessieve erfelijke aandoening β-thalassemie komt in drie verschillende vormen voor: de eerste is homozygote thalassemie, een ernstige ziekte die terug te voeren is op uw onzinmutatie. Heterozygote thalassemie is een mildere ziekte waarbij de nonsensmutaties alleen in een enkele kopie van het β-globine-gen worden aangetroffen. Door het mechanisme van door onzin gemedieerd mRNA-verval kan het mRNA van het defecte gen zodanig worden afgebroken dat alleen gezonde genen tot expressie worden gebracht.
Bij heterozygote thalassemie en dus de matige vorm van de ziekte zit de nonsensmutatie in het laatste mRNA-exon, waardoor de controlemechanismen niet geactiveerd worden. Om deze reden wordt naast gezond β-globine ook verkort β-globine gevormd.Erytrocyten met het defecte β-globine vergaan.
Een ander voorbeeld van het falen van het controlemechanisme is de spierdystrofie van Duchenne, die ook te wijten is aan een onzinmutatie in het mRNA. In dit geval breekt het controlemechanisme het mRNA af, maar veroorzaakt zo een totaal verlies van het zogenaamde dystrofine-eiwit.



.jpg)

.jpg)
.jpg)


.jpg)